КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Розвиток шкіри риб та її похідних
|
|
|
|
Шкіра круглоротих та риб і її похідні
ЛЕКЦІЯ №3
Контакт тварини з зовнішнім середовищем здійснюється за посередництвом ектодерми. Тому сприйняття подразнення, чутливість і захист тіла від шкідливих впливів являють собою основні функції ектодерми. Спеціалізація окремих клітин і цілих ділянок ектодермального покриву для сприйняття тих чи інших подразнень приводить до їх високої диференціації
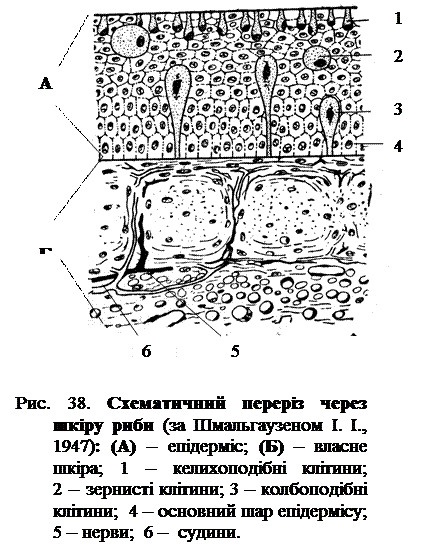 У хребетних тварин завжди чітко помітні два шари шкіри: верхній, ектодермальний – епідерміс і нижній, мезодермальний – дерма або коріум (рис. 38). Епідерміс хребетних завжди має вигляд багатошарового епітелію, нижній шар якого складається з циліндричних клітин. Верхній шар плоских клітин у риб утворює на своїй поверхні тонку пористу облямівку подібно до такої ланцетника. Така тонка облямівка характерна для миготливого епітелію, тому що в личинок ланцетника і молодих ембріонів костистих риб дійсно спостерігається покрив з миготливих війок, то облямівка епідермісу і вважається залишком загального миготливого покриву. Справжньої кутикули, що служить зовнішнім скелетом, у хребетних ніколи не розвивається.
У хребетних тварин завжди чітко помітні два шари шкіри: верхній, ектодермальний – епідерміс і нижній, мезодермальний – дерма або коріум (рис. 38). Епідерміс хребетних завжди має вигляд багатошарового епітелію, нижній шар якого складається з циліндричних клітин. Верхній шар плоских клітин у риб утворює на своїй поверхні тонку пористу облямівку подібно до такої ланцетника. Така тонка облямівка характерна для миготливого епітелію, тому що в личинок ланцетника і молодих ембріонів костистих риб дійсно спостерігається покрив з миготливих війок, то облямівка епідермісу і вважається залишком загального миготливого покриву. Справжньої кутикули, що служить зовнішнім скелетом, у хребетних ніколи не розвивається.
У нижчих хребетних епідерміс відмежований від його сполучної частини шкіри безструктурною основною перетинкою, що є спільним продуктом виділення обох компонентів шкіри. Між клітинами епідермісу прекрасно видно протоплазматичні зв'язки у вигляді міжклітинних містків. По міжклітинних проміжках відбувається циркуляція живильних рідин, що одержують тільки таким шляхом доступ до клітин епідермісу. В епідермісі нижчих хребетних спостерігаються окремі залозисті клітини різних типів, іноді у величезній кількості.
Сполучнотканинна частина шкіри, дерма або коріум хребетних, розвивається за рахунок шкірного листка міотому зародка. У ланцетника цей листок зберігає ще характер епітелію, який виділяє безклітинну опорну тканину драглистого характеру, що знаходиться під епідермісом. У хребетних у цій тканині з'являються і клітинні елементи внаслідок розпаду шкірного листка на окремі клітини – мезенхіму. Однак у личинок нижчих хребетних (круглоротих) мезодермальні клітини розташовуються безперервним шаром, подібно епітелію, під ектодермою, і потім виділяють на своїй поверхні основну речовину із сполучнотканинними волокнами. По мірі росту цього проміжного шару в нього включаються потім і клітинні елементи. У такий спосіб розвивається сполучнотканинна частина шкіри. Волокна шкіри у риб розташовуються дуже правильно – чергуються шарами з взаємно перпендикулярним напрямком волокон, що йдуть під кутом до осі тіла. Шари горизонтальних волокон пронизуються пучками вертикальних волокон, разом з якими в шкіру входять судини і нерви, що її живлять.
В мезодермальній частині шкіри зустрічається пігмент, який концентрується завжди в спеціальних, дуже складно розгалужених пігментних клітинах. Крім звичайних пігментних клітин різних кольорів, розрізняють ще іридоцити, наповнені кристаликами гуаніну, що додають сріблистий блиск багатьом рибам. Пігментні клітини знаходяться у дермі, але вони можуть знаходитись і у міжклітинних проміжках епідермісу.
Рухливість являє собою характерну рису пігментних клітин. Іноді особливо розвинута рухливість пігментних клітин пов'язана із здатністю тварини змінювати своє забарвлення. Такі клітини називаються хроматофори. Зміна забарвлення досягається в цих випадках або переміщенням зерен пігменту з відростків у тіло клітини і навпаки, або скороченням усього хроматофора. Здатність змінювати забарвлення серед риб спостерігається у камбали, скорпени та ін. Пігмент може переміщатися по відростках під самий епідерміс і може пересуватися назад усередину шкіри в тіло клітини. Такі переміщення стимулюються нервами, що підходять до хроматофорів.
Клітини епідермісу виявляють здатність до секреторної діяльності, яка виражається у виділенні їхньою протоплазмою різноманітних рідких речовин, особливо слизу. Такі залозисті клітини розсіяні в епідермісі нижчих хребетних іноді в колосальній кількості, як, наприклад, у круглоротих. У міксин епідерміс складається не менш як на половину усієї своєї товщини винятково з залозистих клітин. І у круглоротих і у риб є залозисті клітини різного типу. Найбільш поширені звичайні келихоподібні слизові клітини, що відкриваються безпосередньо на поверхні епідермісу (рис. 38). Крім того, зустрічаються ще в середині епідермісу кулясті клітини з зернистим вмістом і великі колбоподібні клітини, що сидять на ніжці, яка доходить до основи епідермісу, де вона оточена основним шаром циліндричних клітин. Ці клітини виділяють, очевидно, серозну рідину не безпосередньо на поверхню епідермісу, а в міжклітинні проміжки. Типових багатоклітинних шкірних залоз у риб не зустрічається. Залозисті заглиблення епідермісу біля основи різного роду голок і шипів служать у деяких риб у якості отруто-видільних залоз. Складні, утворення залозистого характеру являють собою й органи світіння деяких глибоководних риб (деякі акули і деякі костисті). У таких органах розрізняють центральну масу клітин залозистого характеру, оточених світло-відбивною сполучнотканинною оболонкою з кристалами гуаніну і покритих з боку, поверненому у середину тіла, пігментною оболонкою. Спереду до залозистої частини органа часто примикають прозорі маси клітин, що грають роль лінзи, а до останнього – ще особливий рефлектор, що складається з увігнутого світло-відбивного дзеркальця, покритого ззаду пігментом.
У шкірі нижчих хребетних майже завжди є тверді кісткові пластинки, які утворють щось подібне до панциру, який захищає тіло тварини. Такі пластинки лежать у сполучнотканинній частині шкіри і називаються луска. У риб зустрічаються три основні види луски, які розрізняються як за формою, так і за матеріалом, з якого вони побудовані. Це – плакоїдна, ганоїдна і кісткова або еласмоїдна луска. Усі ці види луски пов'язані перехідними формами і, представляють результат видозміни одного, найбільш примітивного типу – плакоїдної луски.
 Плакоїдна луска складається з досить широкої основи що знаходиться у власне шкірі (дермі або коріумі) і розташованого на ній шипа, вістря якого висувається через епідерміс назовні. Шип має різну форму: вигнутого назад простого конуса, плоского тризубця тощо.
Плакоїдна луска складається з досить широкої основи що знаходиться у власне шкірі (дермі або коріумі) і розташованого на ній шипа, вістря якого висувається через епідерміс назовні. Шип має різну форму: вигнутого назад простого конуса, плоского тризубця тощо.
Основа найчастіше є у вигляді ромбічної пластинки (рис. 39). Усередині луски є порожнина, заповнена м'якоттю багатою кровоносними судинами, яка називається пульпа (pulpa). Плакоїдна луска складається з дуже твердої речовини – дентину, що представляє з'єднання органічної речовини із солями вапна. Дентин ще твердіший, ніж кістка, і на відміну від останньої, клітинних елементів не містить. Від внутрішньої порожнини в радіальних напрямках відходить безліч тонких розгалужених канальців, що пронизують всю товщу дентину плакоїдної луски. Іноді вершина шипа покрита тонким ковпачком ще більш твердої речовини, у якому немає чітких канальців. Ця речовина містить багато вапна і називається емаль. У канальці дентину заходять відростки клітин, які вистилають внутрішню порожнину. Завдяки утворюючої дії цих клітин, які називаються склеробласти, відбувається наростання маси дентину.
Така луска залягає у шкірі селахій, покриваючи все їхнє тіло дрібними шипиками (рис. 39). На щелепах подібна плакоїдна луска крупніша і має вигляд зубів. Подібність у будові цих органів така велика, що нерідко і звичайну плакоїдну луску називають шкірним зубом. В міру зношування стара плакоїдна луска скидаються (так само, як і зуби на щелепах) і заміняється новою.
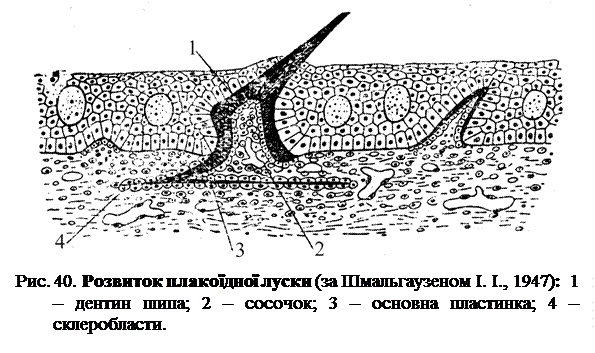 Плакоїдна луска розвивається на межі ектодерми і мезодерми (рис. 40). Нижній шар ектодерми набуває форму ковпачка, у який знизу у вигляді сосочка входить маса мезодермальних клітин. Клітини основного шару епідермісу, що утворюють стінку ковпачка, мають високу циліндричну форму, а мезодермальні клітини, що знаходяться у середині ковпачка розташовуються безпосередньо під епідермісом, та утворюють суцільний шар, що нагадує епітелій і вистилає порожнину ковпачка (рис. 40). Клітини цієї мезодермальної вистилки називаються склеробласти. Перша закладка самої плакоїдної луски виявляється у вигляді потовщення основної перетинки, яка знаходиться під епідермісом. Таким чином, закладка відбувається саме на межі між ектодермою і мезодермою. Шар твердої речовини, який утворюється, являє собою емаль у тому випадку, якщо вона з'являється як результат утворюючої дії основного шару епідермісу. У селахій, однак, частіше зачаток плакоїдної луски з'являється відразу в результаті спільної діяльності основного шару епідермісу і шару мезодермальних склеробластів. Цей початковий шар, що розвивається потім покриває верхівку зуба і дуже твердий; взагалі він дуже подібний до емалі, відрізняється від неї головним чином лише наявністю найтонших канальців. Цю речовину вважають особливим різновидом дентину. У всякому разі цей вітродентин, як його називають, і по своєму складу і по походженню дуже близький до емалі. Подальше наростання речовини в процесі розвитку плакоїдної луски відбувається зсередини і, отже, за рахунок утворюючої дії склеробластів. Останні утворюють на своїй зовнішній поверхні все нові маси дентину, і зростаючий таким чином шип пробивається через епідерміс назовні. Надалі розвиток плакоїдної луски закінчується вже цілком у межах мезодерми, де склеробласти, розташовуються у два шари, між ними виділяють дентинову пластинку, яка служить основою луски. Мезодермальний сосочок виявляється тепер усередині луски, утворюючи його м'якоть. Таким чином, плакоїдна луска стає цілком сформованою.
Плакоїдна луска розвивається на межі ектодерми і мезодерми (рис. 40). Нижній шар ектодерми набуває форму ковпачка, у який знизу у вигляді сосочка входить маса мезодермальних клітин. Клітини основного шару епідермісу, що утворюють стінку ковпачка, мають високу циліндричну форму, а мезодермальні клітини, що знаходяться у середині ковпачка розташовуються безпосередньо під епідермісом, та утворюють суцільний шар, що нагадує епітелій і вистилає порожнину ковпачка (рис. 40). Клітини цієї мезодермальної вистилки називаються склеробласти. Перша закладка самої плакоїдної луски виявляється у вигляді потовщення основної перетинки, яка знаходиться під епідермісом. Таким чином, закладка відбувається саме на межі між ектодермою і мезодермою. Шар твердої речовини, який утворюється, являє собою емаль у тому випадку, якщо вона з'являється як результат утворюючої дії основного шару епідермісу. У селахій, однак, частіше зачаток плакоїдної луски з'являється відразу в результаті спільної діяльності основного шару епідермісу і шару мезодермальних склеробластів. Цей початковий шар, що розвивається потім покриває верхівку зуба і дуже твердий; взагалі він дуже подібний до емалі, відрізняється від неї головним чином лише наявністю найтонших канальців. Цю речовину вважають особливим різновидом дентину. У всякому разі цей вітродентин, як його називають, і по своєму складу і по походженню дуже близький до емалі. Подальше наростання речовини в процесі розвитку плакоїдної луски відбувається зсередини і, отже, за рахунок утворюючої дії склеробластів. Останні утворюють на своїй зовнішній поверхні все нові маси дентину, і зростаючий таким чином шип пробивається через епідерміс назовні. Надалі розвиток плакоїдної луски закінчується вже цілком у межах мезодерми, де склеробласти, розташовуються у два шари, між ними виділяють дентинову пластинку, яка служить основою луски. Мезодермальний сосочок виявляється тепер усередині луски, утворюючи його м'якоть. Таким чином, плакоїдна луска стає цілком сформованою.
Луска селахій складається в основному з дентину, але останній зустрічається в декількох модифікаціях. Емаль, або більш щільний вітродентин, що дуже подібний до емалі, покриває шип у вигляді ковпачка. Головна маса шипа складається з типового дентину, пронизаного тонкими розгалуженими канальцами, у які проникають протоплазматичні відростки склеробластів. Основа плакоїдної луски складається з дентину, що містить складну систему каналів, які є продовженням центральної порожнини і містять також м'якоть із судинами і склеробластами, відростки яких проникають у канальці дентину. Такий дентин схожий на кістку з її гаверсовими каналами і називається трабекулярний дентин, або вазодентин. У зовнішньому шарі основної пластинки знаходяться також характерні для кістки колагенові волокна, що тут не вапнявіють. Таким чином, ця тканина дійсно близька до справжньої кістки. Однак у ній усе-таки зустрічаються в невеликій кількості характерні для дентину канальці, у які проникають відростки склеробластів. Таку тканину, що складає головну масу великої луски ромбових скатів (Raja), називають іноді псевдодентином. Нарешті, іноді дентин приймає шарувату будову і, включаючи клітинні елементи, стає ще більш подібним до кістки.
 У представників кісткових риб Osteіchthyes тіло в більшості випадків покрито лускою, випадки відсутності луски порівняно рідкі. У різних групах кісткових риб зустрічається луска трьох типів: космоїдна, ганоїдна і еласмоїдна. Усі вони є похідними дерми і пов'язані між собою рядом перехідних положень, начебто випливаючи один з одного.
У представників кісткових риб Osteіchthyes тіло в більшості випадків покрито лускою, випадки відсутності луски порівняно рідкі. У різних групах кісткових риб зустрічається луска трьох типів: космоїдна, ганоїдна і еласмоїдна. Усі вони є похідними дерми і пов'язані між собою рядом перехідних положень, начебто випливаючи один з одного.
Два підкласи кісткових риб – лопатнопері Sarcopterygіі і променепері Actіnopterygіі – помітно розрізняються за типом лускатного покриву. У представників лопатноперих луска космоїдна, тоді як у променеперих - у більшості випадків еласмоїдна або ганоїдна і лише далеко не усіх форм (у дводишних Dіpnoі) - космоїдна.
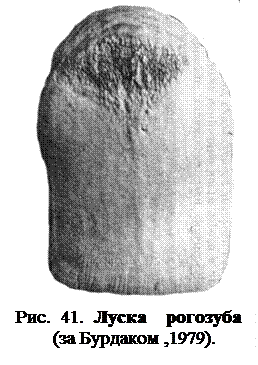
У представників лопатноперих космоїдна луска має ромбічну або округлу форму (рис. 41), луски налягають одна на одну черепицеподібно, утворюючи суцільний лусковий покрив.
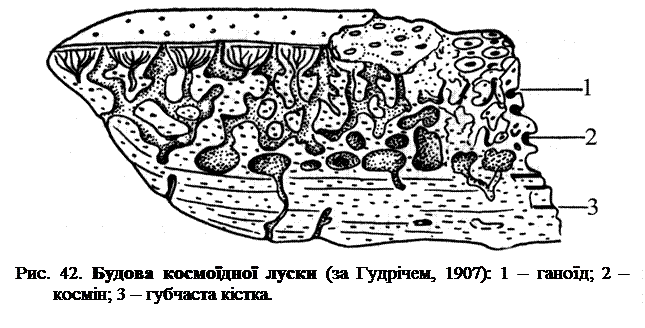 У древніх лопатноперих Crossopterygіі, космоїдна луска складається з декількох шарів, різних за своєю будовою. У її основі знаходиться товста ізопединова базальна пластинка, що містить кісткові клітини і має шарувату будову завдяки горизонтальному орієнтуванню остеобластів. На ізопединовій основі лежить шар губчастої кістки, а на ньому - шар печеристого дентину, який називається косміном. Самий верхній і тонкий шар луски утворений блискучою міцною емаллю - ганоїном - і пронизаний безліччю дрібних отворів, якими відкриваються канали, що йдуть з губчастого космінового шару (рис. 42).
У древніх лопатноперих Crossopterygіі, космоїдна луска складається з декількох шарів, різних за своєю будовою. У її основі знаходиться товста ізопединова базальна пластинка, що містить кісткові клітини і має шарувату будову завдяки горизонтальному орієнтуванню остеобластів. На ізопединовій основі лежить шар губчастої кістки, а на ньому - шар печеристого дентину, який називається косміном. Самий верхній і тонкий шар луски утворений блискучою міцною емаллю - ганоїном - і пронизаний безліччю дрібних отворів, якими відкриваються канали, що йдуть з губчастого космінового шару (рис. 42).
 Філогенетично косміновий шар являє собою результат консолідації комплексу тісно розміщених шкірних зубів, які спочатку утворюють щільний паркет з окремих елементів, а потім зростаються в суцільну дентинову масу з внутрішніми порожнинами, які відповідають пульпарним порожнинам окремих шкірних зубів. Зверху суцільна дентинова маса вкрита консолідованим емалевим покривом. Таким чином, космоїдну луску можна розглядати як похідне луски плакоїдного типу.
Філогенетично косміновий шар являє собою результат консолідації комплексу тісно розміщених шкірних зубів, які спочатку утворюють щільний паркет з окремих елементів, а потім зростаються в суцільну дентинову масу з внутрішніми порожнинами, які відповідають пульпарним порожнинам окремих шкірних зубів. Зверху суцільна дентинова маса вкрита консолідованим емалевим покривом. Таким чином, космоїдну луску можна розглядати як похідне луски плакоїдного типу.
У ході еволюції лопатноперих Sarcopterygіі космоїдная луска поступово видозмінюється за рахунок резорбції космінового шару до окремих горбиків або гребенів, як це спостерігається в деяких ріпідістій і в целокантів, або навіть до повного зникнення косміну, що має місце у сучасних дводишних.
Поверхневий рельєф космоїдної луски, якщо він є, у різних випадках може бути утворений окремими горбиками або ктеноподібними дентиновими зубчиками (латимерія) (рис. 43) або гребенями, орієнтованими уздовж подовжньої осі тіла
Покривні кістки лопатноперих Sarcopterygіі за своєю будовою виявляють значну подібність з космоїдною лускою. Вони складаються з трьох шарів. Нижній шар, як і у космоїдній лусці, представлений товстою ізопединовою пластинкою. На ній лежить середній – найбільш могутній – шар губчастої кістки, пронизаний безліччю судинних каналів. На губчастому шарі знаходиться шар зрослих дентинових шкірних зубів з коронками, що розширюються догори, які утворюють суцільну паркетну поверхню, покриту емалеподібним шаром дуродентину (рис. 44).
 Ганоїдна луска являє собою похідне луски космоїдного типу, відмінності її полягають у поступовій трансформації шарів косміну і губчастої кістки у потовщений верхній ганоїдний шар.
Ганоїдна луска являє собою похідне луски космоїдного типу, відмінності її полягають у поступовій трансформації шарів косміну і губчастої кістки у потовщений верхній ганоїдний шар.
На перших етапах цієї трансформації між ізопединовою основою луски і відносно товстим ганоїновим верхнім шаром ще зберігається тонкий шар косміну з лакунами і каналами (рис. 45). Це так званий палеоніскоїдний підтип ганоїдної луски. Цей же вид ганоїдної луски притаманний сучасним представникам багатоперих (багатопер, каламоіхтес).
Подальша трансформація космінового шару призводить до його повного зникнення. Луска в цьому випадку зберігає лише два шари: нижній - ізопединовий і верхній – ганоїновий (рис. 46). Цей варіант ганоїдної луски називається лепізостеоїдним, оскільки з числа сучасних форм він притаманний Lepіsosteus.
Поверхневий рельєф ганоїдної луски в більшості випадків являє собою сукупність подовжньо орієнтованих гребенів і лунок. Іноді рельєф утворений окремими горбиками або шкірними зубами.
 У ряді випадків шар ганоїну покриває і кістки черепа. У більш пізніх груп луска немає ганоїнового шару і являє собою луску еласмоїдного типу, тобто стає цілком кістковою, представляючи собою тонку кісткову пластинку, що знаходиться у шкірній кишені. Еласмоїдна луска складається з тонких концентрично розташованих кісткових пластинок, що лежать одна на одній на зразок піраміди; сама нижня пластинка – найбільша – є самою молодою, сама верхня – найменша – самою старою. На поверхні утворюючих луску пластинок є характерний рельєф з ребер – склеритів, що виконують функцію ребер жорсткості і одночасно перешкоджають зсуву покриваючого луску епітелію під дією гідродинамічних сил тертя, які виникають при плаванні риби.
У ряді випадків шар ганоїну покриває і кістки черепа. У більш пізніх груп луска немає ганоїнового шару і являє собою луску еласмоїдного типу, тобто стає цілком кістковою, представляючи собою тонку кісткову пластинку, що знаходиться у шкірній кишені. Еласмоїдна луска складається з тонких концентрично розташованих кісткових пластинок, що лежать одна на одній на зразок піраміди; сама нижня пластинка – найбільша – є самою молодою, сама верхня – найменша – самою старою. На поверхні утворюючих луску пластинок є характерний рельєф з ребер – склеритів, що виконують функцію ребер жорсткості і одночасно перешкоджають зсуву покриваючого луску епітелію під дією гідродинамічних сил тертя, які виникають при плаванні риби.
За характером поверхні розрізняють два варіанти еласмоїдної луски: циклоїдну, без зубчиків на вільній поверхні, і ктеноїдну, із зубчиками – ктенодами на вільній поверхні, в іншому обидва ці варіанта цілком ідентичні (рис. 47).
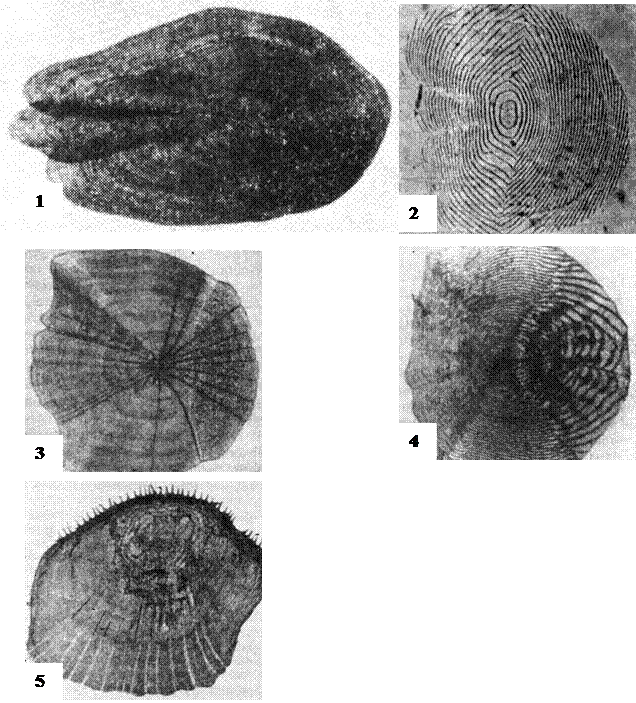
 В онтогенезі і філогенезі риб циклоїдна і ктеноїдна луска за певних умов переходять одна в одну, тобто являють собою дві стадії розвитку того самого пристосування. В онтогенезі ктеноїдна луска проходить циклоїдну стадію. Зовні еласмоїдна луска покрита епідермісом.
В онтогенезі і філогенезі риб циклоїдна і ктеноїдна луска за певних умов переходять одна в одну, тобто являють собою дві стадії розвитку того самого пристосування. В онтогенезі ктеноїдна луска проходить циклоїдну стадію. Зовні еласмоїдна луска покрита епідермісом.
Як правило, еласмоїдна луска утворює суцільний покрив, який характеризується тим, що луски черепицеподібно налягають одна на одну. Рідше луски лежать у шкірі вільно, не торкаючись одна одної (ошибні, зубатки). Порівняно рідкі і випадки часткової або повної редукції еласмоїдного лускатого покриву. Часткова редукція його характерна, наприклад, для деяких скумбрієвих. Повна редукція еласмоїдного лускатого покриву – рідкісне явище і зустрічається найчастіше або в риб, що дуже повільно плавають (соми, гімнеліси, лікоди, пінагори, ліпаріси, морські собачки, і т.п.), або, навпаки, у тих, що дуже швидко плавають (меч-риби та ін.).
Еласмоїдна луска притаманна величезній більшості сучасних променеперих, зокрема, усім костистим рибам (починаючи з оселедцеподібних) і окремим представникам більш примітивних груп (амієві).
Рельєф поверхні еласмоїдної луски складений з елементів, що можуть бути розділені на три категорії.
По-перше, це різні види концентричних рельєфів, утворених краями окремих концентрично розташованих пластинок луски, тобто річними кільцями, а іноді також і склеритами, якщо вони розташовуються концентрично, як, наприклад, у лососевих або щукових (рис. 47, (1)).
По-друге – рельєфи, утворені склеритами не концентричного типу, у вигляді прямих або дугоподібних ліній, орієнтованих у цілому перпендикулярно до напрямку потоку (рис. 47, (2)).
По-третє – сукупність добре обтічних подовжньо орієнтованих елементів поверхневого рельєфу, які у свою чергу можуть бути зведені до двох типів: каналами стоку, утворених поздовжніми зниженнями поверхні з більш-менш плавним поперечним перерізом (рис. 47, (3)), і ктеноподібними структурами, представленими більш конкретними утвореннями – окремими горбками (рис. 47, (4)), зубцями і подовжніми колами з більш різким поперечним перерізом, або більш довгими поздовжньо спрямованими шипами, що розташовані на вільному краї луски і налягають на відкрите поле наступної луски (рис. 47, (5)).
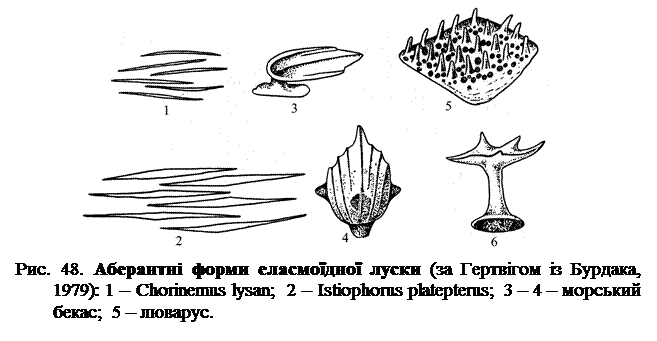 У ряду представників променеперих еласмоїдна луска зустрічається в дуже зміненій, аберрантній формі (рис. 48.). Як приклади можна вказати подовжену, голкоподібну луску деяких ставридок (рис. 48 (1 – 2)), дуже своєрідну луску морського бекаса із пластинчастою, розташованою на ніжці коронкою (рис. 48 (3 – 4)) озброєну сильними вертикальними зубцями луски спинорогів та ін. (рис. 48 (5)), луску люваруса з грибоподібною зубчастою коронкою (рис. 48 (6)), і деякі інші. Усі ці незвичайні форми луски в кожному конкретному випадку мають своє функціональне значення.
У ряду представників променеперих еласмоїдна луска зустрічається в дуже зміненій, аберрантній формі (рис. 48.). Як приклади можна вказати подовжену, голкоподібну луску деяких ставридок (рис. 48 (1 – 2)), дуже своєрідну луску морського бекаса із пластинчастою, розташованою на ніжці коронкою (рис. 48 (3 – 4)) озброєну сильними вертикальними зубцями луски спинорогів та ін. (рис. 48 (5)), луску люваруса з грибоподібною зубчастою коронкою (рис. 48 (6)), і деякі інші. Усі ці незвичайні форми луски в кожному конкретному випадку мають своє функціональне значення.
В деяких випадках у променеперих, коли еласмоїдна луска відсутня, тіло покрите кістковими пластинками, що має місце, зокрема, у морських лисичок, підкаменщиків, пучкозябрових, колюшкоподібних; у представників останньої родини ці пластинки утворюють суцільний панцир.
У макроморфології еласмоїдного лускового покриву виділяються дві основні особливості, характерні для більшості представників костистих: черепицеподібне налягання луски і рядність її розташування на тілі риби.
Черепицеподібне налягання луски забезпечує лусковому покриву певну міцність і, у той же час, майже не утрудняє локомоторні згинання тіла риби.
Розташування луски в лусковому покриві поздовжніми рядами (рядність луски) забезпечує оптимальні умови обтікання тіла риби. У типовому випадку розташування рядів луски на тілі збігається з лініями току води, що обтікає рибу.
|
|
|
|
Дата добавления: 2014-01-05; Просмотров: 8322; Нарушение авторских прав?; Мы поможем в написании вашей работы!