КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Потенциал покоя в клетках
|
|
|
|
Финал
Итак:
Калий-натриевый насос создает предпосылки для возникновения потенциала покоя. Это - разность в концентрации ионов между внутренней и наружной средой клетки. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка клетки выравнять концентрацию ионов по калию приводит к потере калия, потере положительных зарядов и порождает электроотрицательность внутри клетки. Эта электроотрицательность составляет большую часть потенциала покоя. Меньшую его часть составляет непосредственная электрогенность ионного насоса, т.е. преобладающие потери натрия при его обмене на калий.
Потенциал покоя - стационарная разность электрических потенциалов, между внутренней и наружной поверхностями мембраны в невозбужденном состоянии.
Потенциал покоя определяется разной концентрацией ионов по обе стороны мембраны и диффузией ионов через мембрану.
Если концентрация иона внутри клетки Свн отлична от концентрации этого иона снаружи Снар и мембрана проницаема для этого иона, возникает поток заряженных частиц через мембрану, вследствие чего нарушается электрическая нейтральность системы, образуется разность потенциалов внутри и снаружи клетки  , которая будет препятствовать дальнейшему перемещению ионов через мембрану. При установлении равновесия выравниваются значения электрохимических потенциалов по разные стороны мембраны:
, которая будет препятствовать дальнейшему перемещению ионов через мембрану. При установлении равновесия выравниваются значения электрохимических потенциалов по разные стороны мембраны: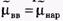 Так как
Так как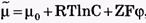 , то
, то
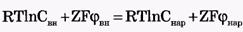
Отсюда легко получить формулу Нернста для равновесного мембранного потенциала:
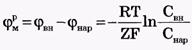
Если мембранный потенциал обусловлен переносом ионов К+, для которого 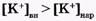 и Z = +1, равновесный мембранный потенциал
и Z = +1, равновесный мембранный потенциал
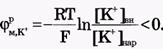
Для ионов Na+:[Na+]вн<[Na+]нap, Z = +1,
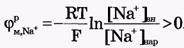
Для ионов Сl-:[Сl-]вн<[Сl-]нар, Z = -1 и
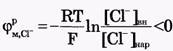
Если в формуле Нернста перейти от натурального логарифма к десятичному, то для положительного одновалентного иона (Z = +l)
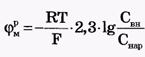
Примем температуру Т = 300 К, тогда
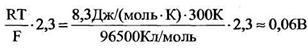
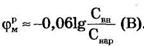
Согласно Бернштейну (1902 г.), причина мембранного потенциала покоя - диффузия ионов калия из клетки наружу.
Примем в формуле Нернста Cвн/Снар≈100, что по порядку величины соответствуют экспериментальным данным для калия 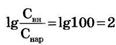 и мембранный потенциал
и мембранный потенциал 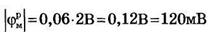 , что несколько больше модуля экспериментально измеренных значений потенциала покоя.
, что несколько больше модуля экспериментально измеренных значений потенциала покоя.
Рассчитано что изменение концентрации ионов в клетке вследствие выхода из клетки 10-17 моль ионов составит 2*10-3 ммоль/л. Это ничтожное изменение концентрации по сравнению с изменением концентрации ионов калия внутри клетки, составляет всего 10-4 % от концентрации калия внутри клетки. Таким образом, чтобы создать равновесный нернстовский мембранный потенциал, через мембрану должно пройти пренебрежимо малое количество ионов по сравнению с общим их количеством в клетке.
В табл. 1 приведены значения мембранного потенциала, рассчитанного по формуле Нернста для различных клеток и для различных ионов, и экспериментально полученные значения потенциала покоя для этих клеток.
Таблица 1. Содержание ионов К+, Na+, C1-, равновесные потенциалы и потенциалы покоя некоторых клеток.
| Объект | Концентрация, ммоль/л | мB по формуле Нернстаf | мВ эксперf | |||||||||
| [К+] | [Na+] | [С1] | К+ | Na+ | С1- | |||||||
| вн. | нар. | вн. | нар. | вн. | нар. | |||||||
| Гигантский аксон | 360 | 10 | 70 | 420 | 160 | 500 | -90 | +50 | -30 | -60 | ||
| Мышца лягушки | 125 | 2,5 | 15 | 125 | 11 | 120 | -98 | +60 | -87 | -94 | ||
Из сравнения рассчитанных и экспериментальных значений мембранного потенциала видно, что потенциал покоя на самом деле ближе к потенциалу, рассчитанному по формуле Нернста для К+. Вместе с тем, обращает на себя внимание значительное расхождение экспериментальных и теоретических значений. Причины расхождения в том, что не учтена проницаемость мембраны для других ионов. Одновременная диффузия через мембрану ионов К+, Na+ и С1- учитывается уравнением Гольдмана. Уравнение Гольдмана можно вывести из уравнения Нернста-Планка. В стационарном случае, когда, возникая на мембране, разность потенциалов - мембранный потенциал - тормозит дальнейший перенос ионов через мембрану, суммарный поток различных ионов становится равным нулю: 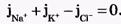 Перед Jcl стоит знак минус, учитывающий отрицательный заряд иона хлора. Однако, так как в создании мембранного потенциала участвуют различные ионы, равновесие при этом не наступает, потоки различных ионов не равны нулю по отдельности. Если учесть только потоки JK+ и JNa+, то Перед Jcl стоит знак минус, учитывающий отрицательный заряд иона хлора. Однако, так как в создании мембранного потенциала участвуют различные ионы, равновесие при этом не наступает, потоки различных ионов не равны нулю по отдельности. Если учесть только потоки JK+ и JNa+, то 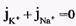 , или , или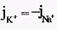 и получим: и получим: 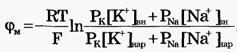 Если учесть еще и поток ионов С1-, то можно получить уравнение для мембранного потенциала, созданного потоками через мембрану трех видов ионов, уравнение Гольдмана: Если учесть еще и поток ионов С1-, то можно получить уравнение для мембранного потенциала, созданного потоками через мембрану трех видов ионов, уравнение Гольдмана: 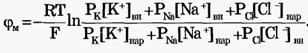 В числителе выражения, стоящего под знаком логарифма, представлены концентрации [К+]ВН, [Na+]BH, но [С1-]НАР, а в знаменателе - [К+]НАР, [Na+]НАР, но [С1-]ВН, так как ионы хлора отрицательно заряжены. В состоянии покоя проницаемость мембраны для ионов К+ значительно больше, чем для Na+, и больше, чем для С1-: РК» РNa, РК > Р С1 Для аксона кальмара, например, РK:PNa:PC1=1:0,04:0,45 Переписав уравнение Гольдмана в виде: В числителе выражения, стоящего под знаком логарифма, представлены концентрации [К+]ВН, [Na+]BH, но [С1-]НАР, а в знаменателе - [К+]НАР, [Na+]НАР, но [С1-]ВН, так как ионы хлора отрицательно заряжены. В состоянии покоя проницаемость мембраны для ионов К+ значительно больше, чем для Na+, и больше, чем для С1-: РК» РNa, РК > Р С1 Для аксона кальмара, например, РK:PNa:PC1=1:0,04:0,45 Переписав уравнение Гольдмана в виде: 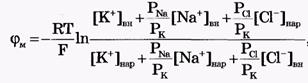 в случае, когда проницаемость мембраны для ионов натрия и хлора значительно меньше проницаемости для калия: РNa «РК и РС1 «РК из уравнения Гольдмана получим уравнение Нернста для мембранного потенциала покоя: в случае, когда проницаемость мембраны для ионов натрия и хлора значительно меньше проницаемости для калия: РNa «РК и РС1 «РК из уравнения Гольдмана получим уравнение Нернста для мембранного потенциала покоя: 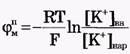 Таким образом, уравнение Нернста - частный случай уравнения Гольдмана. Мембранный потенциал, рассчитанный по уравнению Гольдмана, оказался по абсолютной величине меньше мембранного потенциала, рассчитанного по формуле Нернста, ближе к экспериментальным его значениям в крупных клетках. И формула Нернста, и уравнение Гольдмана не учитывают активного транспорта ионов через мембрану, наличия в мембранах электрогенных (вызывающих разделение зарядов, а следовательно и возникновение разности потенциалов) ионных насосов, играющих важную роль в поддержании ионного равновесия в мелких клетках. В цитоплазматической мембране работают K+-Na+-ATФaзы, перекачивающие калий внутрь клетки, а натрий из клетки. С учетом работы электрогенных ионных насосов для мембранного потенциала было получено уравнение Томаса (Томас, 1972 г.): Таким образом, уравнение Нернста - частный случай уравнения Гольдмана. Мембранный потенциал, рассчитанный по уравнению Гольдмана, оказался по абсолютной величине меньше мембранного потенциала, рассчитанного по формуле Нернста, ближе к экспериментальным его значениям в крупных клетках. И формула Нернста, и уравнение Гольдмана не учитывают активного транспорта ионов через мембрану, наличия в мембранах электрогенных (вызывающих разделение зарядов, а следовательно и возникновение разности потенциалов) ионных насосов, играющих важную роль в поддержании ионного равновесия в мелких клетках. В цитоплазматической мембране работают K+-Na+-ATФaзы, перекачивающие калий внутрь клетки, а натрий из клетки. С учетом работы электрогенных ионных насосов для мембранного потенциала было получено уравнение Томаса (Томас, 1972 г.): 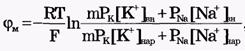 где m - отношение количества ионов натрия к количеству ионов калия, перекачиваемых ионными насосами через мембрану. Чаще всего K+-Na+-ATФaза работает в режиме, когда m =3/2, m всегда больше 1. (Нет ионных насосов, перекачивающих С1-, поэтому в уравнении Томаса отсутствуют члены РС1 [С1-].) Коэффициент m>1 усиливает вклад градиента концентрации калия в создание мембранного потенциала, поэтому мембранный потенциал, рассчитанный по Томасу, больше по абсолютной величине, чем мембранный потенциал, рассчитанный по Гольману, и дает совпадение с экспериментальными значениями для мелких клеток. Нарушение биоэнергетических процессов в клетке и работы K+-Na+-ATФазы приводит к уменьшению |φм|, в этом случае мембранный потенциал лучше описывается уравнением Гольд-мана. Повреждение клеточной мембраны приводит к повышению проницаемости клеточных мембран для всех ионов: к повышению и РК, и PNa, и РС1. Вследствие уменьшение различия проницаемостей абсолютное значение мембранного потенциала |φм| снижается. Для сильно поврежденных клеток |φм| еще меньше, но сохраняется отрицательный мембранный потенциал φм за счет содержащихся в клетке полианионов - отрицательно заряженных белков, нуклеиновых кислот и других крупных молекул, не могущих проникнуть через мембрану (доннановский потенциал). где m - отношение количества ионов натрия к количеству ионов калия, перекачиваемых ионными насосами через мембрану. Чаще всего K+-Na+-ATФaза работает в режиме, когда m =3/2, m всегда больше 1. (Нет ионных насосов, перекачивающих С1-, поэтому в уравнении Томаса отсутствуют члены РС1 [С1-].) Коэффициент m>1 усиливает вклад градиента концентрации калия в создание мембранного потенциала, поэтому мембранный потенциал, рассчитанный по Томасу, больше по абсолютной величине, чем мембранный потенциал, рассчитанный по Гольману, и дает совпадение с экспериментальными значениями для мелких клеток. Нарушение биоэнергетических процессов в клетке и работы K+-Na+-ATФазы приводит к уменьшению |φм|, в этом случае мембранный потенциал лучше описывается уравнением Гольд-мана. Повреждение клеточной мембраны приводит к повышению проницаемости клеточных мембран для всех ионов: к повышению и РК, и PNa, и РС1. Вследствие уменьшение различия проницаемостей абсолютное значение мембранного потенциала |φм| снижается. Для сильно поврежденных клеток |φм| еще меньше, но сохраняется отрицательный мембранный потенциал φм за счет содержащихся в клетке полианионов - отрицательно заряженных белков, нуклеиновых кислот и других крупных молекул, не могущих проникнуть через мембрану (доннановский потенциал).
| ||||||||||||
|
|
|
|
|
Дата добавления: 2014-10-23; Просмотров: 1486; Нарушение авторских прав?; Мы поможем в написании вашей работы!